 Ikke-tilfeldig variasjon
Ikke-tilfeldig variasjon
(fritt oversatt etter boka: 'Not By Chance' av Dr. Lee Spetner kap. 7A; Judaica Press, 1999)
Når tilhengere av neo-darwinistisk teori (NDT) hevder at genetisk variasjon er tilfeldig, innebærer det at de mener at sannsynligheten for at en variasjon inntreffer ikke har noe med måten det hjelper organismen å tilpasse seg til omgivelsene på. Når forfatteren her (Dr. Lee Spetner) hevder at variasjon ikke er tilfeldig, mener han at sjansen for at den inntreffer har noe med hvordan organismen tilpasser seg omgivelsene. Han mener at tilpasningen på noe vis er influert av omgivelsene eller av organismens behov.
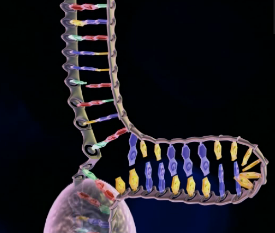
Bilde 1. Balanse mellom seleksjon og variasjon
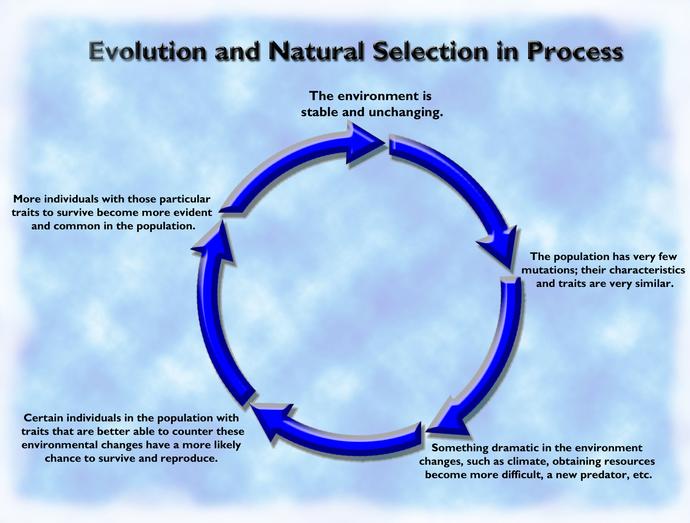
Helt siden formuleringen av NDT, på 1940-tallet, har forskere stilt seg skeptiske til hvilken rolle naturlig seleksjon hadde. Den berømte biologen D'Arcy W. Thompson (1942) kritiserte rollen til naturlig seleksjon: .."den viktige funksjonen til naturlig seleksjon er ikke å være opphav til, men å fjerne.. vi ser i naturlig seleksjon en ubønnhørlig kraft hvis effekt ikke er å skape, men å ødelegge -å luke, beskjære, å kutte ned og kaste i ilden .. (ss269-270)
NDT svarer at kraften til naturlig seleksjon ligger i dens evne til å velge ut. Millioner av nye former blir presentert for den ut fra tilfeldig variasjon. Den velger fra de som er tilpassede. Men vi har tidligere argumentert statistisk for at NDT ikke har evnen til å skape nok informasjon, til å bli selektert. Tidligere har flere biologer og studenter av evolusjon nådd fram til samme konklusjon. Mae-Wan Ho fra Open University of Milton Keynes og Peter Saunders fra universitetet i London, har uttalt: (Ho og Saunders, 1979): -variasjon i fenotypen, som naturlig seleksjon kan virke på, oppstår ikke tilfeldig.. Tilfeldige mutasjoner for å bygge informasjon, er blitt beskrevet som 'ikke-bevisbare' og 'statistisk usannsynlige' Cook (1977) -som stemmer med konklusjonen vi kom til i kap 4.
 NDT som baserer seg på tilfeldige variasjoner, kan ikke gjøre rede for store evolusjonære endringer. Den kan bare gjøre rede for en begrenset gruppe av små endringer. Det er ikke bevis for at tilfeldige mutasjoner og naturlig seleksjon spilte noen rolle i opphavet til noen av hovedgruppene av dyr, inkl. arter (Ho og Saunders, 1979). NDT redegjør ikke for opprinnelsen til dyre-rekkene, klassene eller ordnene. Den begrunner ikke engang artenes opprinnelse, unntatt i spesielle tilfeller som vi kan kalle trivielle, sammenlignet med livets store strøm. Det er ikke dokumentert at disse kan utvides til å gjelde makroevolusjon, f.eks. at ei flue endrer seg til f.eks. veps.
NDT som baserer seg på tilfeldige variasjoner, kan ikke gjøre rede for store evolusjonære endringer. Den kan bare gjøre rede for en begrenset gruppe av små endringer. Det er ikke bevis for at tilfeldige mutasjoner og naturlig seleksjon spilte noen rolle i opphavet til noen av hovedgruppene av dyr, inkl. arter (Ho og Saunders, 1979). NDT redegjør ikke for opprinnelsen til dyre-rekkene, klassene eller ordnene. Den begrunner ikke engang artenes opprinnelse, unntatt i spesielle tilfeller som vi kan kalle trivielle, sammenlignet med livets store strøm. Det er ikke dokumentert at disse kan utvides til å gjelde makroevolusjon, f.eks. at ei flue endrer seg til f.eks. veps.
De dramatiske påstandene knyttet til NDT er det som har påkalt verdens oppmerksomhet mot evolusjon. Prominente evolusjonister har uttrykt meninger i vitenskapens navn, som går utover hva de kan dokumentere vitenskapelig. Den ledende paleonotologen G. G. Simpson har uttalt at 'Darwins teori har gitt mennesket et nytt begrep om seg selv' (Simpson, 1960
). Det har fått den prominente genetiker Bentley Glass til å prøve å utvikle etiske verdier for menneskeheten fra NDT (Glass, 1965).
Bilde 2. Mikro og makro-evolusjon
NDT hevder å kunne gjøre rede for hvordan alt liv utviklet seg fra en enkel begynnelse. Den hevder at utviklingen av livet er et naturlig fenomen. Siden vi ikke kjenner til noen fysiske lover som får organismer til å utvikle seg, foreslo Darwin at det skjedde statistisk i forhold til populasjoner. Men det NDT kan gjøre rede for, er bare noen få isolerte endringer. NDT forklarer ikke hvordan livets informasjon kan ha blitt bygd opp. Når NDT skal prøve å forklare store endringer, så blir argumentasjonen deres bare et scenario av noe som kunne ha skjedd. Opprinnelig da NDT oppsto, så ble makroevolusjon beskrevet som resultatet av en lang rekke små endringer Huxley (1943). Beskrivelsen deres fulgte av Darwins hypotese om akkumulerende seleksjon.
 I starten var det en minoritet av evolusjonister som nektet å akseptere en gradvis prosess som forklaring til makrovevolusjon. De kunne ikke se noen måte hvorpå kumulative seleksjon kunne oppnå den hærskare av tilpasninger, som måtte skje i forbindelse med store evolusjonære endringer. Paleontologen O.H. Schindelwolf (1950) så reelle gap i forsillmaterialet, og han kunne ikke godta en sakte gradvis evolusjon som dets forklaring. Genetikeren Richard Goldsmith (1940, 1952) foreslo å gjøre rede for store evolusjonære endringer som resultat av plutselige, store mutasjoner eller evolusjonære sprang. Zoologen H. Graham Cannon (1958) foreslo en Lamackisk mekanisme kunne virke og at tilpassede variasjoner faktisk kunne bli fremkalt av omgivelsene. Han foreslo imidlertid ikke en mekanisme for å gjøre rede for hvordan dette kunne skje. Men flertallet innen NDT lot seg ikke overtale av minoritetens argumenter. De avviste store mutasjoner som en måte å oppnå store evolusjonære steg på. De helte seg til små mutasjoner, som var kjent for å framkalle mikroevolusjon, til også å begrunne makroevolusjon (Rench 1954, 1960; Simpson 1953). De tok avstand fra større mutasjoner som grunnlag for evolusjon, fordi de nesten alltid er skadelige. Store tilpassede mutasjoner er i prinsippet for sjeldne til å kunne føre til evolusjonære endringer, selv i store populasjoner over lange tidsperioder.
I starten var det en minoritet av evolusjonister som nektet å akseptere en gradvis prosess som forklaring til makrovevolusjon. De kunne ikke se noen måte hvorpå kumulative seleksjon kunne oppnå den hærskare av tilpasninger, som måtte skje i forbindelse med store evolusjonære endringer. Paleontologen O.H. Schindelwolf (1950) så reelle gap i forsillmaterialet, og han kunne ikke godta en sakte gradvis evolusjon som dets forklaring. Genetikeren Richard Goldsmith (1940, 1952) foreslo å gjøre rede for store evolusjonære endringer som resultat av plutselige, store mutasjoner eller evolusjonære sprang. Zoologen H. Graham Cannon (1958) foreslo en Lamackisk mekanisme kunne virke og at tilpassede variasjoner faktisk kunne bli fremkalt av omgivelsene. Han foreslo imidlertid ikke en mekanisme for å gjøre rede for hvordan dette kunne skje. Men flertallet innen NDT lot seg ikke overtale av minoritetens argumenter. De avviste store mutasjoner som en måte å oppnå store evolusjonære steg på. De helte seg til små mutasjoner, som var kjent for å framkalle mikroevolusjon, til også å begrunne makroevolusjon (Rench 1954, 1960; Simpson 1953). De tok avstand fra større mutasjoner som grunnlag for evolusjon, fordi de nesten alltid er skadelige. Store tilpassede mutasjoner er i prinsippet for sjeldne til å kunne føre til evolusjonære endringer, selv i store populasjoner over lange tidsperioder.
Bilde 3. Eks. på mutasjon
De fleste evolusjonister i dag holder variasjon som evolusjon baseres på for å være tilfeldig. Men de er delt i hvordan en kan begrunne makroevolusjon. Majoriteten er tilhengere av små gradvise endringer. De mener at storskala-endringer, som er hva Darwin forsøkte å forklare, utgjøres av lange strenger av mindre endringer. Men de er ikke i stand til å gjøre rede for at det positiv seleksjonsverdig kan inntreffe ofte nok til at naturlig seleksjon har tilstrekkelig å arbeide på, eller informasjonsoppbyggingen som må ha skjedd i levende organismer.
 Tilhengerne av små gradvise endringer må vise at i gjennomsnitt, må kumulativ seleksjon øke litt informasjon i genomet for hvert evolusjonært steg. Men siden genetikk startet som vitenskap, er det ikke funnet en eneste mutasjon som øker informasjon. I prinsippet er det ikke utenkelige, men det er statistisk usannsynlig. Selv om NDT var en attraktiv teori, bygde den på falske forutsetninger om at mange små tilfeldige mutasjoner kunne bygge opp til store evolusjonære endringer. Det var ingen bevis for det i Darwins tid, og det har ikke dukket opp noe bevis siden publikasjonen av Darwins bok inntil NDT ble bearbeidet på 1940-tallet.
Tilhengerne av små gradvise endringer må vise at i gjennomsnitt, må kumulativ seleksjon øke litt informasjon i genomet for hvert evolusjonært steg. Men siden genetikk startet som vitenskap, er det ikke funnet en eneste mutasjon som øker informasjon. I prinsippet er det ikke utenkelige, men det er statistisk usannsynlig. Selv om NDT var en attraktiv teori, bygde den på falske forutsetninger om at mange små tilfeldige mutasjoner kunne bygge opp til store evolusjonære endringer. Det var ingen bevis for det i Darwins tid, og det har ikke dukket opp noe bevis siden publikasjonen av Darwins bok inntil NDT ble bearbeidet på 1940-tallet.
Spekulasjonen ble likevel akseptert som mulig, ja endog som fakta. Men i løpet av perioden NDT har eksistert, så er det ingen av data som rettferdiggjør spekulasjonen i NDT om at stor-skala evolusjon har sin kilde i tilfeldige variasjon. Alle bevis er mot det. De som er tilhengere av sprangvis evolusjon, benekter at den lange, sakte prosessen av små endringer kan fremkalle store evolusjonære endringer. Det kan, i følge deres syn, ha funnet sted ved genetiske rearrangeringer i regulerende gener. Bare en endring i timing av et molekylært signal under embryo-fasen, kunne resultere i større endringer i fenotypen (Gould 1977).
Bilde 4. Hundens 'utvikling'
 De fleste tilhengere av sprangvis evolusjon støtter tilfeldig utvikling (randomness). Men om utviklingen er tilfeldig, så må de også kunne redegjøre for oppbygging av informasjon. Sjansen er bortimot null for at en større mengde informasjon kan ha kommet inn i genomet på det viset. Det er derfor majoriteten innen NDT støtter sin teori på en lang rekkefølge av suksessive små-skala endringer. Etter at dette har vist seg vanskelig, har noen tilhengere av teorien forsøkt å redde teorien ved å fortolke variasjonene som større endringer i regulerende gener. De holder likevel fast på naturlige, tilfeldige endringer. Men en kan ikke forvente å få større, tilpassede endringer ved tilfeldighet. Inne i cellen synes genetiske rearrangeringer å være en normal aktivitet, som styres av spesifikke enzymer, som cellen produserer akkurat for det formålet -så vidt vi kjenner til. Noen biologer foretrekker å kalle dem tilfeldige, bare fordi deres timing ennå ikke er forstått.
De fleste tilhengere av sprangvis evolusjon støtter tilfeldig utvikling (randomness). Men om utviklingen er tilfeldig, så må de også kunne redegjøre for oppbygging av informasjon. Sjansen er bortimot null for at en større mengde informasjon kan ha kommet inn i genomet på det viset. Det er derfor majoriteten innen NDT støtter sin teori på en lang rekkefølge av suksessive små-skala endringer. Etter at dette har vist seg vanskelig, har noen tilhengere av teorien forsøkt å redde teorien ved å fortolke variasjonene som større endringer i regulerende gener. De holder likevel fast på naturlige, tilfeldige endringer. Men en kan ikke forvente å få større, tilpassede endringer ved tilfeldighet. Inne i cellen synes genetiske rearrangeringer å være en normal aktivitet, som styres av spesifikke enzymer, som cellen produserer akkurat for det formålet -så vidt vi kjenner til. Noen biologer foretrekker å kalle dem tilfeldige, bare fordi deres timing ennå ikke er forstått.
Bilde 5. Miljømessige faktorer påvirker utvikling
Noen ønsker å si at en organisme kan øke sin informasjon ved å assimilere et stykke DNA fra en annen organisme (horisontal genoverføring). Det kan forekomme innenfor bakterier, men om vi fokuserer på hvordan informasjon bygges opp, så må vi adressere spørsmålet hvordan informasjonen oppsto i den andre organismen. Å forsøke å bygge informasjon ved å overføre informasjon fra art til art, er som å forsøke å bygge en økonomi ved å la alle ta inn andres klesvask. For at en økonomi skal vokse, så må den bli skapt. For at informasjon skal øke i levende organismer, så må den dannes ett sted.
Om variasjon som fører til stor-skala endringer, ikke er tilfeldig, så må vi sjekke om ikke-tilfeldige kilder for variasjon kan være årsak til det. Observerte tilpasninger frembragt av omgivelsene er blitt rapportert i langt over hundre år, men er i hovedsak blitt ignorert av dominerende biologi -grunnet dominans av NDT. Slike observasjoner er gjerne feid under teppet, fordi de ikke kunne forstås innen rammeverket av teorien. Noen genetikere har følt seg presset til å unnskylde for observasjoner av dette slaget. De betraktet dem som 'uheldige defekter i det delikate genetiske maskineriet (West-Eberhard 1989). I de siste tre tiårene av det 20. århundre begynte biologer å foreslå at variasjonen som drev evolusjon kanskje ikke var tilfeldig (Ho og Saunders 1979; Cook 1977; Rosen og Butch 1980). Variasjonen ble trigget av omgivelsene, og organismen tilpasset seg dem. Cook ( 1977) bemerket at NDT sto på svakt grunnlag både matematisk og biokjemisk, og han uttrykte seriøs tvil omkring nytten av tilfeldige variasjone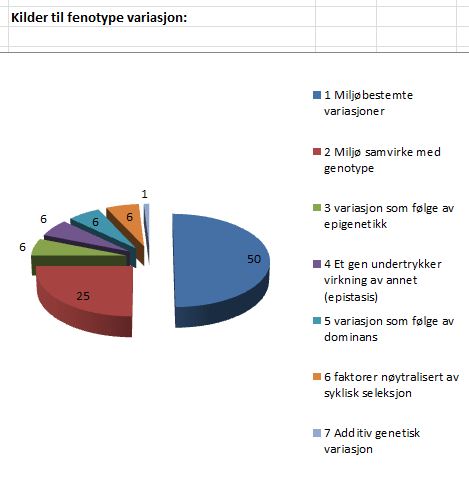 r i evolusjon. Bare en minoritet av biologer holder på at evolusjonens variasjon ikke kan være tilfeldig, men i dette tilfellet er minoriteten på mer solid grunn enn majoriteten. En er nødt til å vurdere ikke-tilfeldig variasjon om en ønsker sann innsikt om emnet evolusjon.
r i evolusjon. Bare en minoritet av biologer holder på at evolusjonens variasjon ikke kan være tilfeldig, men i dette tilfellet er minoriteten på mer solid grunn enn majoriteten. En er nødt til å vurdere ikke-tilfeldig variasjon om en ønsker sann innsikt om emnet evolusjon.
Levende organismer responderer på miljøet på ulike måter. For å overleve og klare seg bra, senser en organisme omgivelsene og tilpasser seg etter det. I genetisk kontroll av enzymer i cellen, senser det genetiske kontrollapparatet nærheten av enzymets substrat, og slår PÅ genet som koder for enzymet. Systemet slår gener AV eller PÅ, etter som de trengs, men det skjer ingen permanente endringer i genomet. Den slags kontrollmekanismer tillater organismen å operere effektivt under ulike kortvarige endringer som miljøet kan variere innenfor. En utvidelse av slike kontroller kan føre til endringer på langvarig evolusjonært nivå. Om kontrollen er i utviklingsprosessen, så kan små endringer av 'rett slag' føre til store adaptive tilpasninger av fenotypen. Men slike endringer av 'rett slag' vil ikke være sannsynlige å inntreffe bare ved tilfeldighet. Om de skulle være tilfeldige, ville sannsynligheten for en 'riktig endring' være proporsjonal med andelen av 'rette endringer' blant alle mulige endringer. Antall endringer som fører til feil, er enormt mye større enn 'rette' endringer. Men om genomet er satt til å svare på signal fra omgivelsene, så ville ikke tilfeldigheter være involvert. En 'riktig' tilpasset endring ville sikkert inntreffe når den trengtes.
Det er ulike typer variasjon av fenotypen, som kan iverksettes av miljøet, og flere av dem kan føre til langvarige endringer i en populasjon. Variasjoner kan deles i to ulike hovedgrupper: I den første er variasjonen i fenotypen som resulterer fra endringer i DNA-sekvensen. I den andre gruppen er variasjoner i fenotypen, uten endringer i DNA-sekvensen. Ikke-tilfeldige mutasjoner hører til den første gruppen.. Mutasjoner er endringer i DNA-sekvensen som kan deles i ulike typer. Noen er endring i enkeltnukleotider, og noen er mer komplekse. Mange mutasjoner er kjent for å være spontane, og effekten er uavhengig av miljøets innflytelse. Det er ikke, etter forfatterens mening, mutasjoner som leder til makroevolusjon. Bevis indikerer at disse mutasjonene er resultatet av feil i genetisk virkemåte, hvorav de fleste er kopieringsfeil i DNA. Mutasjonene som forfatteren er mest opptatt av, er de som viser tegn på å være ikke-tilfeldige, ved at de trigges av miljøet omkring organismen. De tilhører den første klassen av mutasjoner som kan føre til observert evolusjon. Disse kan fungere som svitsjer som trigges av omgivelsene. Disse mutasjonene kan svitsje genomet til ett av et sett eksisterende potensielle tilstander, for å produsere en tilpasset fenotype. Vi har sett at tilfeldige mutasjoner ikke øker informasjon i genomet: NDT forutsetter at en lang kjede av tilfeldige endringer kan føre til store evolusjonære endringer. Men med alle eksperimentelle resultat i genetikk som er kjent for forfatteren, finnes det ikke bevis for en viktig forutsetning i NDT: 'En tilpasning som øker genomets informasjon kan alltid finne sted gjennom å endre en enkelt nukleotide'. Denne forutsetningen er et essensielt punkt i deres teori. Mangel på bevis for dette punktet gjør at vi må avvise forutsetningen, og dermed hele NDT. At det ikke er påvist mutasjoner som øker genomets informasjon, viser at det slett ikke finnes 'millioner på millioner av slike mutasjoner' som teorien forutsetter. Kanskje er det ingen? At man ikke har lykkes med å finne en eneste mutasjon med økt informasjonsinnhold, innebærer mer enn manglende støtte, det er bevis mot teorien. Mutasjoner som bidrar til varige endringer er ikke-tilfeldige. De er trigget av omgivelsene og gir tilpassede fenotyper. Potensialet for tilpasning i forhold til miljøet, er allerede til stede i genomet. Omgivelsene bare trigger eksiterende muligheter.
Bilde 6. Kilder til fenotype variasjon (fra Kimura s.30-31)
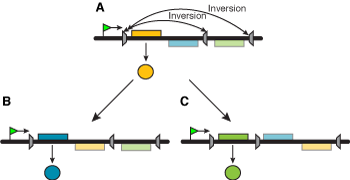 Ikke alle mutasjoner er tilfeldige. Noen er kjent for å være ikke-tilfeldige i tid, andre i sted. Noen inntreffer akkurat når de trengs, og noen inntreffer akkurat på rett sted. Mange mutasjoner (se senere) er utført med presisjon og er under utstrakt cellulær kontroll. Å påstå at de er tilfeldige, vil være å vende det døve øret til hva naturen vil si oss. Forfatterens hentydninger her er 'spekulative': i) Vi har ennå ikke (1997) et eks. på én mutasjon som er tilstrekkelig godt forstått til at vi kan se hvordan omgivelsene trigger den, og ii) hvordan det produseres en fenotype som er tilpasset miljøet. Vi vet at trigging inntreffer, men ikke hvordan. En velkjent type mutasjon som ikke synes å være helt tilfeldig, er en inversjon av en DNA-seksjon. En inversjon innen et gen, kan skape problemer for genet, og gjøre det inaktivt. Om inversjonen på et senere tidspunkt blir presist reversert, så vil genet gjenvinne sin aktivitet. Evt. kan en inversjon fjerne en aktivator fra en regulerende gensekvens, og slå AV hele komplekset av relaterte gener. Presise inversjoner kan således tjene enten til å stoppe eller restarte genetisk aktivitet. Inversjoner kan derved produsere plutselige, store endringer i fenotypen Darnell et al. (1986). Inversjonsmønstre i kromosomer har vært benyttet til å karakterisere noen arter av bananfluen (Drosophila -Dobzhansky 1951). Det er mulig at inversjonene i seg selv var genetiske endringer som førte til dannelse av en ny art bananflue.
Ikke alle mutasjoner er tilfeldige. Noen er kjent for å være ikke-tilfeldige i tid, andre i sted. Noen inntreffer akkurat når de trengs, og noen inntreffer akkurat på rett sted. Mange mutasjoner (se senere) er utført med presisjon og er under utstrakt cellulær kontroll. Å påstå at de er tilfeldige, vil være å vende det døve øret til hva naturen vil si oss. Forfatterens hentydninger her er 'spekulative': i) Vi har ennå ikke (1997) et eks. på én mutasjon som er tilstrekkelig godt forstått til at vi kan se hvordan omgivelsene trigger den, og ii) hvordan det produseres en fenotype som er tilpasset miljøet. Vi vet at trigging inntreffer, men ikke hvordan. En velkjent type mutasjon som ikke synes å være helt tilfeldig, er en inversjon av en DNA-seksjon. En inversjon innen et gen, kan skape problemer for genet, og gjøre det inaktivt. Om inversjonen på et senere tidspunkt blir presist reversert, så vil genet gjenvinne sin aktivitet. Evt. kan en inversjon fjerne en aktivator fra en regulerende gensekvens, og slå AV hele komplekset av relaterte gener. Presise inversjoner kan således tjene enten til å stoppe eller restarte genetisk aktivitet. Inversjoner kan derved produsere plutselige, store endringer i fenotypen Darnell et al. (1986). Inversjonsmønstre i kromosomer har vært benyttet til å karakterisere noen arter av bananfluen (Drosophila -Dobzhansky 1951). Det er mulig at inversjonene i seg selv var genetiske endringer som førte til dannelse av en ny art bananflue.
 Bilde 1. Eks. på gen-inversjon
Bilde 1. Eks. på gen-inversjon
Ett interessant eks. på en inversjon er en som ofte opptrer i bakterien Salmonella. Denne mutasjonen resulterer i erstatning av en type protein med en annen (Silverman og Simon 1983, Darnell 1986 ss441-442; Griffits et al 1993 ss.572-573). Disse to proteinene er så vidt vi vet identiske i funksjon. De skiller seg bare ad ved sine antistoff-egenskaper. Genet som koder for disse proteinene kan være i én av to tilstander. I en tilstand, blir ett av proteinene lagd i den andre det andre proteinet. En spesifikk og presis inversjon er det som gjør at genomet går fra den ene til den andre tilstanden. Ett segment av genomet på ca. 1000 nukleotider synes å bli spesifikt tilordnet til inversjonen. Ved hver ende har segmentet spesielt korte DNA-sekvenser på 14 nukleotider, som tillater inversjonen å inntreffe gjennom en rekombinasjon. Et nødvendig protein for rekombinasjonen blir kodet innen 1000 nukleotiders-seksjonen. De spesielle markeringene ved enden sikrer at segmentet vil bli presist invertert og reinvertert. Inversjonen fungerer som en bryter for å endre flagellin-proteinet. Så langt vi vet, skjer disse inversjonene mer eller mindre regelmessig. De inntreffer omtrent hver 10. generasjon. Periodisk svitsjing fra ett anti-gen til ett annet hjelper bakterien å unngå vertscellens immunsystem. Genomets tilstand er arvelig. Antigenet som ikke blir angrepet av immunsystemet synes å øke i antall. Rekombinasjoner i genomet kan også produsere andre slag av mutasjoner. Det kan frambringe slettinger, dupliseringer og forflytninger. Rekombinasjon er ikke en tilfeldig prosess. Den er under streng genetisk kontroll, og krever atskillige spesifikke enzymer for å kunne utføres. Det finnes t.o.m. spesielle gener som påvirker effekten ved rekombinasjonen (Grittiths og al 1993, s. 571 ff)
Bilde 2. Salmonella bakterien
Det er oppdaget at genetiske elementer flytter fra plass til plass på ett kromosom. De flytter også fra ett kromosom til et annet. Det er snakk om korte stykker av DNA, funnet i bakterier, planter og insekter som kalles flyttbare genetiske elementer. Genetikere tror at disse elementene finnes i alle organismer. Det er gjenkjent to slag av slike, hvorav det minste har opp mot 1.500 nukleotider, som kalles innsettings (insertion)-sekvenser (IS). De større kalles for transposoner, har mer enn 2.500 nukleotider -noen har så mange som 20.000. Så vidt vi vet kan innsettingssekvenser ta plass i hvilken som helst del av genomet. Om de kommer inni genet, forvrenger de det genet og gjør det ineffektivt. Et IS kan inneholde transkripsjon-terminerings signaler, som i så fall slår AV hele strengen av lenkede gener (operon) som signalstrømmen følger innen genet det havner inni. Den samme innsettingssekvensen (IS) er kommet til syne i ulike organismer, og har fått et tall lagt til som navn: f.eks. IS1 og IS5. Tilsvarende kalles de større transposonene for TnX, der X står for ett siffer, f.eks. Tn1 og Tn903. Et transposon som har resistens mot ampicillin (Tn1) har ca. 5.000 nukleotider. Transposonet som bærer resistens mot tetracycline (Tn10) har ca. 10.000 nukleotider. Transposonet som innehar resistens mot ampicillin, streptomycin og sulfanilamide (Tn4) har 20.000 nukleotider. De kan flytte fra et arvestoff i bakterien inn i dens DNA. {Det kan også skje horisontal genoverføring, fra en bakterie til en annen -oversetters anmerkning.}
Stoffutvalg og bilder ved Asbjørn E. Lund samt {kommentert -i klammeparentes}
.